
Ostatnia aktualizacja 28 lipca 2022
Związki te działają bez wiedzy ich nadawcy i odbiorcy w świecie ludzi i zwierząt w tym najliczniejszej ich grupy stawonogów (owadów). Przez kilka ostatnich lat istnieje trend (głównie u kobiet) kupowania flakoników z rzekomo zawartością tych związków w swoim składzie. Panie kupują te specyfiki dla zdobycia “upatrzonego” partnera lub odzyskania jego względów.
Mowa będzie o związkach semiochemicznych. Badania potwierdzają wpływ związków semiochemicznych na zachowanie ludzi:
- androstadienon wzmacnia chęć do współpracy pomiędzy mężczyznami przy podejmowaniu decyzji
- androstadienon sygnalizuje męskość heteroseksualnym mężczyznom, a estratetraenol sygnalizuje kobiecość heteroseksualnym kobietom
- kobiety częściej nawiązują kontakt wzrokowy z mężczyzną, na którego zaaplikowano mieszaninę androsteronu i androstenolu
- androstadienon zmienia atrakcyjność mężczyzn w oczach kobiet
- wąchanie androstadienonu i estratetraenolu wpływa na nastrój, pamięć oraz funkcjonowanie autonomicznego układu nerwowego w specyficznych kontekstach
- związki semiochemiczne kobiet karmiących piersią zwiększają motywację seksualną
- androstadienon podświadomie skupia zasoby psychiczne człowieka na bodźcach o znaczeniu emocjonalnym
- dodatek hormonów płciowych do perfum ma właściwości atraktanta seksualnego zarówno dla mężczyzn, jak i kobiet
- wąchanie androstadienonu wpływa pozytywnie na nastrój i pobudzenie
- kobietom, które zostały poddane działaniu androstadienonu, zwiększa się poziom kortyzolu w ślinie
Do najbardziej znanych związków semiochemicznych należą FEROMONY, ale o nich póżniej.
Jakie czynniki wpływają na to, że ktoś nam się podoba?
Składniki atrakcyjności “fizycznej”
- Twarz: kształt twarzy, wielkość, położenie i kształt jej elementów, stan skóry
- Sylwetka: obwód w pasie, bioder, klatki piersiowej, wielkość piersi
- Wysokość ciała, długość nóg
- Ręka (dłoń): kształt, skóra
- Głos: wysokość
- Zapach
- Ruch: głowy, części twarzy (np. mimika), chód, taniec
Składniki atrakcyjności “niefizycznej”
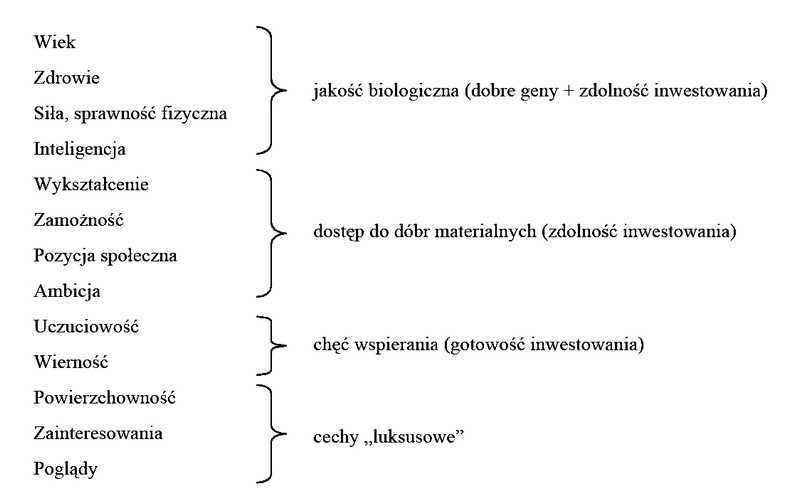
- Wiek
- Zdrowie jakość biologiczna (dobre geny + zdolność inwestowania) ; Siła, sprawność fizyczna
- Inteligencja
- Wykształcenie
- Zamożność dostęp do dóbr materialnych (zdolność inwestowania); Pozycja społeczna; Ambicja
- Uczuciowość chęć wspierania (gotowość inwestowania)
- Wierność
- Powierzchowność
- Zainteresowania cechy „luksusowe”
- Poglądy
Dymorfizm płciowy wymagań u człowieka i nie tylko
(użyte symbole ♀ kobieta ♂ mężczyzna )
♀ inwestuje w potomstwo więcej niż ♂ (ciąża, laktacja, opieka)
♀ może dużo stracić, jeśli skojarzy się z „niewłaściwym” ♂
(np. wadliwy materiał genetyczny ♂, niechęć ♂ do opieki)
♀ jest bardziej wybiórcza (wybredna) przy szukaniu partnera niż ♂
Dymorfizm płciowy preferencji partnerskich
| Ważniejsze dla ♂ niż dla ♀ | Ważniejsze dla ♀ niż dla ♂ |
| – płodność: młodość, atrakcyjność fizyczna – wierność fizyczna | – zdolność i gotowość inwestowania – wierność psychiczna – dobre geny (gł. do związku krótkotrwałego) |
Korzyści pośrednie i bezpośrednie
Korzyści bezpośrednie (= materialne): oddziaływania fenotypu partnera, np. opieka, ochrona.
Korzyści pośrednie (= genetyczne): dobre geny partnera przekazywane potomstwu.
Dobroć genów ma 2 składniki:
– addytywny: wynika z wartości genotypowej allela (dobroć „sama w sobie”)
– nie-addytywny: zależy od obecności innych genów (dominacja, epistaza).
Ważne jest więc dopasowanie genów ojca i matki – komplementarność genów.
Wartość partnerska
Osobnik X ma dla osobnika Y wysoką wartość partnerską =
= związek z osobnikiem X jest dla osobnika Y korzystny z reprodukcyjnego punktu widzenia.
Wysoka wartość partnerska – warto się z nim związać
Niska wartość partnerska – nie warto się z nim wiązać
Dany osobnik może mieć wysoką wartość partnerską dla osoby A, a niską dla osoby B – jeżeli osoby A i B mają inne potrzeby (i preferencje).
Dany osobnik może mieć inną wartość partnerską w kontekście związku krótkotrwałego niż długotrwałego (dobre geny vs dobra osobowość).
Dobór płciowy u zwierząt
♀ inwestuje w potomstwo więcej niż ♂
♀ jest bardziej wybredna niż ♂
atrakcyjność fizyczna partnera jest ważniejsza dla ♀ niż dla ♂ czyli odwrotnie niż u człowieka – dlaczego?
U wielu gatunków ♀ może od ♂ uzyskać tylko korzyści pośrednie (dobre geny)
♀ preferuje u ♂ takie oznaki dobrych genów jak: symetria, przesadne cechy, żywe kolory
U niektórych gatunków ♂ dostarcza ♀ korzyści bezpośrednich
♀ ceni oznaki zdolności inwestowania u ♂ (np. karmienie godowe u ptaków)
Gdy ♂ może dostarczać korzyści genetycznych i materialnych to optymalną strategią dla ♀ jest:
- utworzyć związek długotrwały z ♂ chętnym do inwestowania w potomstwo
- potajemnie kojarzyć się z ♂ o dobrych genach
Skutek: partner długotrwały przykładnie opiekuje się nie swoim potomstwem o dobrych genach.
Samcowi o dobrych genach opłaca się być kochankiem wielu samic – nie traci czasu na opiekę.
Samcowi o gorszych genach pozostaje zostać dobrym ojcem (i mężem).
Koewolucja preferencji i preferowanych cech
Co było pierwsze: preferencja czy cecha? Obie możliwości dozwolone:
- Istnieje preferencja dla jakiegoś bodźca (skutek sposobu funkcjonowania układu nerwowego) –
– u płci przeciwnej powstaje preferowana cecha –
– najlepsze biologicznie osobniki potrafią lepiej wytworzyć tę cechę –
– cecha ta staje się oznaką jakości biologicznej –
– nasila się preferencja dla tej cechy
- Pewna cecha pomaga w przeżyciu lub w konkurencji wewnątrzpłciowej –
– najlepsze biologicznie osobniki potrafią lepiej wytworzyć tę cechę –
– cecha ta jest oznaką jakości biologicznej –
– pojawia się preferencja dla tej cechy
Atrakcyjność twarzy – młodzieńczość
Kobieta – młody wygląd (proporcje nastolatki) – oznaka płodności
Mężczyzna – średni wiek (obie skrajności obniżają atrakcyjność) – oznaka dojrzałości i sprawności Dziecko – młody wygląd („niemowlęce” proporcje) – czy jest oznaką czegoś?
Osoby starsze – stosunkowo młody wygląd – oznaka zdrowia
Atrakcyjność twarzy – przeciętność proporcji
Proporcjonalność = przeciętność proporcji = typowość proporcji =
= każdy wymiar twarzy jest równy średniej populacyjnej.
Przeciętność jest ważna u ♀ i ♂, z przodu i z profilu.
Wygląd proporcjonalnej twarzy jest różny w różnych populacjach (rasach).
Przeciętność jest oznaką jakości genetycznej.
Atrakcyjność twarzy – cechy płciowe
Nasilenie cech płciowych = „typowość płciowa” = maskulinizacja u ♂ i feminizacja u ♀.
Preferowane są:
- silnie sfeminizowane (kobiece) twarze ♀ – oznaka młodości i płodności; nieznacznie zmaskulinizowane (męskie) twarze ♂
♂ o silnie zmaskulinizowanych twarzach (tacy są i tak są postrzegani):
- dobre geny i wysoka jakość biologiczna
- niedobry charakter
♀ idą na kompromis, wybierając ♂ o jakimś stopniu maskulinizacji.
Dlaczego opozycja dobre geny-dobry charakter występuje u ♂, ale nie u ♀?
Atrakcyjność twarzy – symetria
Lewa strona twarzy jest lustrzanym odbiciem prawej strony.
Preferowany jest znaczny stopień symetrii, ale nie idealna symetria.
Symetria jest oznaką jakości genetycznej.
Atrakcyjność twarzy – inne komponenty
Ogólnie: oznaki młodości i kobiecości u ♀, a umiarkowana maskulinizacja u ♂.
| ♀ | ♂ | |
| Kolor skóry (i włosów) | jasny | raczej ciemny |
| Zarost twarzy | brak | „szczecinka” |
| Stan skóry | bez zmarszczek i skaz | to samo, ale nie tak ważne |
| Uśmiech | istotny | nieistotny |
| Wielkość źrenic | duża | umiarkowana |
Atrakcyjność twarzy statyczna (na zdjęciu) silnie koreluje z dynamiczną (film na żywo).
Samoocena atrakcyjności twarzy słabo koreluje z ocenami osób postronnych.
Wewnątrzpopulacyjne zróżnicowanie preferencji
Matematycznie rzecz ujmując, stopień międzyosobniczej zgodności w postrzeganiu atrakcyjności twarzy jest równie znaczny jak stopień międzyosobniczej rozbieżności.
Czyli, udział indywidualnego gustu w ocenie atrakcyjności twarzy wynosi 50%.
Ponadto, istnieje znaczna niezgodność sędziego samego z sobą – gdy powtórnie ocenia atrakcyjność grupy twarzy, to ocenia je inaczej niż za pierwszym razem.
Wewnątrzpopulacyjne zróżnicowanie preferencji – wiek
Atrakcyjność fizyczna i seksowny wygląd jest ważny dla osób młodych.
Niemowlęta i noworodki mają preferencje podobne do dorosłych.
Wewnątrzpopulacyjne zróżnicowanie preferencji – płeć
Twarze ♀ są atrakcyjniejsze wg ♂ niż wg ♀.
Twarze ♂ są atrakcyjniejsze wg ♀ niż wg ♂.
Twarze ♀ są atrakcyjniejsze niż twarze ♂ wg ♀ i ♂.
Wewnątrzpopulacyjne zróżnicowanie preferencji – jakość biologiczna
Osoby zdrowe, silnie dymorficzne, atrakcyjne fizycznie i uważające się za atrakcyjne:
- przykładają dużą wagę do atrakcyjności fizycznej partnera
- preferują oznaki jakości biologicznej na twarzach osób płci przeciwnej:
- dymorfizm
- symetria
- zdrowy wygląd
Wewnątrzpopulacyjne zróżnicowanie preferencji – stan fizjologiczny
- ♀ w fazie płodnej cyklu miesiączkowego – silnie preferuje dymorficzne i symetryczne twarze ♂
- Ciężarna ♀ – słabo preferuje seksowne twarze ♂
- ♂ o wysokim poziomie testosteronu – preferuje silnie sfeminizowane twarze ♀
Wewnątrzpopulacyjne zróżnicowanie preferencji – osobowość i stan psychiczny
- Kto szuka osoby ekstrawertycznej / asertywnej / ciepłej, ten preferuje twarze sugerujące ekstrawertyzm / asertywność / ciepło.
- ♂ w dobrym nastroju woli ♀ atrakcyjną niż miłą; przygnębiony ♂ – odwrotnie.
Wewnątrzpopulacyjne zróżnicowanie preferencji – warunki oceny
- Tłok lub gorące pomieszczenie – niższe oceny atrakcyjności.
- Podprogowa ilość feromonu – wyższe oceny atrakcyjności.
Wewnątrzpopulacyjne zróżnicowanie preferencji – sytuacja życiowa
- Osoba jest w stałym związku, mniej jej się podobają atrakcyjne osoby płci przeciwnej.
- ♂ jest w trudnej sytuacji materialnej, preferuje twarze ♀ sugerujące siłę fizyczną i psychiczną.
Wewnątrzpopulacyjne zróżnicowanie preferencji – podobieństwo twarzy
♂ i ♀ preferują twarze podobne do twarzy własnej.
Dokładniej rzecz ujmując:
♂ i ♀ preferują twarze podobne do twarzy rodzica płci przeciwnej.
Podobieństwo dotyczy:
- proporcji twarzy
- koloru włosów i oczu
- rasy partnera (kolor skóry)
- wieku partnera (młodzieńczości wyglądu twarzy)
Preferencji takich brak, jeżeli w dzieciństwie miało się złe relacje z rodzicem płci przeciwnej.
Wewnątrzpopulacyjne zróżnicowanie preferencji – uprzednio widziane twarze
- Podobają się nam twarze, które:
- prędzej oglądaliśmy (efekt czystej ekspozycji)
- podobne do tych, które prędzej oglądaliśmy (uogólniony efekt ekspozycji)
- Oglądanie atrakcyjnych twarzy, kolejne twarze wydają się mniej atrakcyjne (efekt kontrastu)
- Twarze oglądane w niemiłych warunkach (dzwonek), wydają się mniej atrakcyjne przy powtórnym oglądaniu w normalnych warunkach (warunkowanie).
Wewnątrzpopulacyjne zróżnicowanie preferencji – znajomość i wiedza
- Twarze osób znajomych bardziej się nam podobają niż osób nieznajomych.
- Twarze lubianych znajomych bardziej się podobają niż nie lubianych.
- W szczęśliwych związkach, twarz partnera wydaje się atrakcyjniejsza niż jest naprawdę. W nieudanych związkach jest odwrotnie.
- Informacje (pozytywne / negatywne), jakie posiadamy o osobie, wpływają (pozytywnie / negatywnie) na ocenę atrakcyjności jej twarzy.
Międzypopulacyjne podobieństwo preferencji
- Niektóre oznaki pożądanych cech są uniwersalne, np. gładka skóra – młodość.
- Globalizacja kultury zachodniej – upowszechnianie zachodnich standardów atrakcyjności.
Międzypopulacyjne zróżnicowanie preferencji
- Preferencja dla przeciętnych twarzy jest uniwersalna, ale przeciętna twarz ma różny wygląd w różnych populacjach.
- Różne populacje żyją w różnych w różnych warunkach, a preferencje są zależne od warunków.
Korelacje pomiędzy atrakcyjnością różnych cech ciała
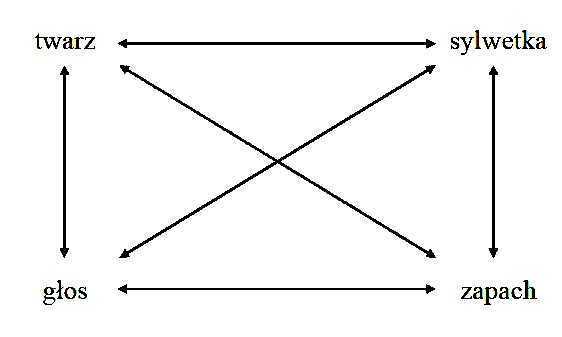
Dotyczy to ♂ oraz ♀ i wynika m. in. z poziomu hormonów płciowych (estrogenu i testosteronu).
Korelacje pomiędzy preferencjami dla różnych cech ciała
Preferencje ♂ dla ♀:
- sfeminizowane twarze zapach żeńskich feromonów
Preferencje ♀ dla ♂:
- zapach męskich feromonów
- zmaskulinizowane twarze
- niski, męski głos
Neurofizjologiczne reakcje na widok atrakcyjnych twarzy
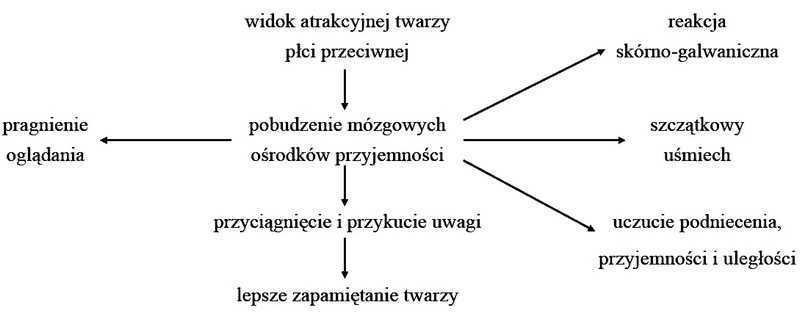
Atrakcyjność twarzy a wartość partnerska
Cechy osób o atrakcyjnych twarzach:
- geny zapewniające lepszą odporność na pasożyty
- rzadsze choroby infekcyjne
- dłuższe życie
- większa sprawność fizyczna
- wyższy iloraz inteligencji
- lepsze zdrowie psychiczne
- wyższe stężenia hormonów płciowych – większa płodność
- ruchliwsze i zwykle prawidłowo zbudowane plemniki (♂)
Konsekwencje biologiczne atrakcyjności
Cechy osób o atrakcyjnych twarzach:
- popularność „randkowa”
- atrakcyjność seksualna
- ♂ doprowadzają ♀ do orgazmu
- więcej partnerów
- wcześniej ślub ♀
- rzadziej stara panna
- więcej potomstwa
- większy odsetek córek
Istnieje korelacja atrakcyjności między partnerami.
Konsekwencje społeczne atrakcyjności
- Ocenianie:
- inteligencja, zdrowie, towarzyskość, przystosowanie społeczne
- egoizm, próżność, buta, niewierność
- Traktowanie: faworyzowanie w kontaktach przypadkowych, w pracy, w sądzie, przy głosowaniu.
- Samospełniające się proroctwo: atrakcyjni stają się przystosowani społecznie.
- Samoobalające się proroctwo: nieatrakcyjni zdobywają lepsze wykształcenie i większą kontrolę nad życiem.
- Makijaż, kosmetyki, medycyna estetyczna, chirurgia plastyczna.
Atrakcyjna sylwetka
Preferencje dla sylwetki są silnie zróżnicowane międzypopulacyjnie i w czasie.
We współczesnej kulturze zachodniej:
| atrakcyjna ♀: | atrakcyjny ♂: |
| wyraźne wcięcie w talii (WHR = 0,7) | znaczna wysokość ciała |
| szczupłość (BMI < 20) | mezomorfik (kształt V) |
| piersi przeciętnej wielkości lub większe | symetria ciała |
| długie nogi | długie nogi |
| wysoki głos | niski głos |
Atrakcyjność zapachu
Zapach potu jest atrakcyjny, gdy pochodzi od:
- ♀ w płodnej fazie cyklu miesiączkowego
- osoby heterozygotycznej
- osoby o innych genach na odporność niż osoba oceniająca
Natura preferencji
Preferencje dla ludzkich cech fizycznych:
- w znacznym stopniu są ewolucyjnymi adaptacjami
- częściowo wynikają z tendencyjności percepcyjnej (sposobu działania układu nerwowego)
- w stosunkowo niewielkim stopniu są wynikiem mody
Filogeneza preferencji
Preferencje odziedziczone po małpich przodkach:
- przeciętność
- symetria
- stan skóry
Ludzkie nabytki ewolucyjne:
- preferencja dla młodego wyglądu ♀
- rezerwa wobec znacznej maskulinizacji ♂
Język wzajemnego komunikowania się zwierząt( w tym ssaków) przez wiele lat stanowił temat badań i dyskusji. Podejrzewano, że oprócz sygnałów wizualnych, dotykowych czy dźwiękowych (o których było wyżej) istnieje jeszcze niewidzialna dla oka komunikacja chemiczna, wywierająca znaczący wpływ na życie zwierząt, w szczególności na procesy reprodukcji, zdobywanie pożywienia oraz reakcje obrony i ucieczki. Związki sygnalne biorące udział w regulacji behawioru różnych organizmów zostały nazwane substancjami semiochemicznymi (grec. semio, sygnał).
Wśród nich na szczególną uwagę z całą pewnością zasługują związki infochemiczne, do których zalicza się feromony. W odróżnieniu od hormonów, które oddziałują na inne komórki i tkanki tego samego organizmu, feromony to chemiczne sygnały komunikacji pomiędzy różnymi organizmami. Nazwa „feromon” wywodzi się z języka greckiego, w którym pherein oznacza przekazywać, a hormao — pobudzać. Określenie to zostało po raz pierwszy użyte w 1959 r., kiedy to Karlson i Lüscher zdefiniowali feromony jako substancje wydzielane przez osobniki tego samego gatunku, prowokujące specyficzne reakcje fizjologiczne oraz behawioralne. Definicja ta wymaga jednak pewnego uaktualnienia, ponieważ znane są przykłady międzygatunkowych kontaktów pod wpływem identycznego feromonu, na przykład płciowego. Związki o charakterze feromonów zostały odkryte niespełna pół wieku temu. Obok tych niewątpliwie najbardziej znanych substancji semiochemicznych jakimi są feromony, do tej samej grupy związków zalicza się również:
— autotoksyny — odchody i metabolity toksyczne zarówno dla wytwarzającego je organizmu, jak i dla innych osobników );
— autoinhibitory populacji ograniczające jej nadmierny rozrost powyżej poziomu zakłócającego równowagę ekologiczną;
— nekromony — substancje rozpoznawania śmierci, które wydzielane przez ciała martwych zwierząt odstraszają osobniki tego samego lub innego gatunku.
Istnienie feromonów potwierdzały już obserwacje sprzed wielu stuleci. W XVII w. zauważono, iż pojedyncza pszczoła w czasie ukłucia wydziela substancję zachęcającą inne owady do ataku na uprzednio użądlonego osobnika. Jak się później okazało, najważniejszym składnikiem tej mieszaniny był octan 3-metylobutylu, opisany w 1962 r.. Badania profesora Adolfa Butenandta przyczyniły się do zidentyfikowania w 1959 r. pierwszego feromonu — bombykolu. Związek ten, wydzielany z gruczołów samic jedwabnika morwowego, potrafi zwabić samce z odległości przekraczającej 11 km. Samiec może wyczuć ten zapach w minimalnym jego stężeniu, nawet przy bardzo dużym rozcieńczeniu. Jak dotąd, potwierdzono obecność feromonów u ponad 1500 gatunków zwierząt, z czego zdecydowaną większość stanowią owady.
Odkrycie bombykolu rozpoczęło badania nad budową feromonów. Przez wiele lat twierdzono, że są to substancje jednoskładnikowe. Ostatnie eksperymenty wskazują jednak na fakt, iż feromony to związki wieloskładnikowe, z zazwyczaj jednym dominującym składnikiem. Pod względem stanu skupienia mogą to być substancje lotne lub nielotne. Skład chemiczny związków infochemicznych jest zmienny i zależy od funkcji przekazywanego sygnału, osobnika będącego źródłem feromonu, pory roku czy dnia. Ten sam sygnał, może mieć bardzo wiele odmiennych znaczeń w zależności od warunków środowiskowych. Znaczna część feromonów zawiera w swojej budowie centra chiralne, generujące dużą grupę stereoizomerów, z których zazwyczaj tylko jeden jest biologicznie czynny. Badania chemików pozwoliły na określenie aktywności biologicznej ponad 200 chiralnych feromonów oraz podzielenia tych związków na grupy, w zależności od ich działania oraz przestrzennej konformacji. Do I grupy, najliczniej reprezentowanej, należą związki, spośród których tylko jeden z enancjomerów jest biologicznie czynny. W II grupie spotykamy zależność inhibicji funkcjonowania jednego ze stereoizomerów przez drugi, a III grupa reprezentuje synergistyczne działanie enancjomerów, w którym to nieaktywny lub mało aktywny stereoziomer działa synergistycznie na czynny biologicznie składnik feromonu. W związku z tym, ich wspólne działanie przewyższa sumę aktywności pojedynczych enancjomerów.
Feromony wytwarzane są głównie w narządach wydzielania zewnętrznego. Mogą je stanowić gruczoły o prostej budowie, utworzone z komórek nabłonkowych, czy też wysoce wyspecjalizowane gruczoły, zbudowane z komórek wydzielniczych i kanałowych. Emisja feromonów odbywa się również dwiema drogami. Mogą być one bezpośrednio uwalniane na zewnątrz lub uprzednio gromadzone w specjalnie wytworzonych do tego celu zbiorniczkach, które w zależności od gatunku, feromonu czy pory roku osiągają rozmaite rozmiary.
Po wydzieleniu do środowiska zewnętrznego, feromony, zależnie od struktury chemicznej, docierają do odbiorcy wraz z powietrzem, wodą lub w wyniku bezpośredniego kontaktu. Zasięg oraz szybkość rozprzestrzeniania się tych związków semiochemicznych są zróżnicowane i uzależnione od pełnio nych funkcji. Dlatego też promień działania niektórych feromonów ścieżkowych jest bardzo mały, natomiast feromony płciowe mogą być rozpoznawane z dużej odległości. Różny jest także czas trwania ich aktywności. Przykładem mogą być feromony alarmowe, które, jak sama nazwa wskazuje, cechuje krótkotrwałe, lecz szybkie działanie.
Aby mogło dojść do „semiochemicznej komunikacji” pomiędzy organizmami, feromony muszą dotrzeć do swoistych receptorów na powierzchni komórek odbiorcy. U owadów odbywa się to za pośrednictwem włosków olfaktorycznych (właściwych narządów węchu) występujących na powierzchni anten (czułek). Feromony zaadsorbowane na ich powierzchni dyfundują do wnętrza, ulegają rozpuszczeniu w znajdującym się tam płynie i docierają do receptorów. Receptory takie tworzą siedem domen transbłonowych, które na skutek związania z feromonami aktywują, za pośrednictwem białka G, szlaki sygnalizacji wewnątrzkomórkowej. Komórki receptorowe przekształcają w ten sposób sygnał chemiczny w elektryczne impulsy, które następnie przekazywane są neuronami do płata antenowego (centrum nerwowego owada).
U kręgowców za odbiór feromonów odpowiedzialny jest głównie narząd lemieszowo-nosowy (ang. vomeronasal organ, VNO), zwany również narządem Jacobsona, gdyż to on opisał go po raz pierwszy w 1813 r.. VNO to parzysty organ, który jest umiejscowiony po obu stronach przegrody nosowej. Łącząc się w ten sposób z jamą nosową, odbiera informacje niesione w postaci lotnych związków chemicznych. U wielu zwierząt narząd przylemieszowy może być także połączony z jamą gębową, dzięki czemu służy również jako detektor substancji nielotnych.
VNO to podwójny kanalik chrząstkowy o długości od 2 do 20 cm. W narządzie Jacobsona można wyróżnić trzy części: ślepo zakończoną część ogonową, część pośrednią i głowową, która otwiera się do kanału łączącego jamę nosową z ustną (przewodu siekaczowego). Taka morfologia cechuje VNO większości zwierząt gospodarskich, chociaż istnieją pewne różnice gatunkowe dotyczące szczegółowej budowy. Wyjątkiem są konie, u których odcinkiem ślepo zakończonym jest część głowowa. Jama omawianego narządu wyścielona jest nabłonkiem zmysłowym zbudowanym z komórek receptorowych, podporowych i podstawnych oraz drugim typem nabłonka, utworzonym wyłącznie z komórek podporowych i podstawnych. Pod warstwą nabłonka występuje silnie unaczyniona tkanka jamista. Natomiast grzbietowo i brzusznie w stosunku do jamy VNO znajdują się liczne gruczoły egzokrynne (gruczoły lemieszowe) z niewielkimi kanałami wydzielniczymi uchodzącymi do jamy VNO. Narząd przylemieszowy jest bardzo dobrze unaczyniony. Krew dostarczana jest tętnicą, a odprowadzana jedną lub dwoma żyłami biegnącymi wzdłuż organu w tkance jamistej. VNO jest także silnie unerwiony, zarówno włóknami nerwów współczulnych, jak i przywspółczulnych.
Zwierzęta badają pyskiem określone wydzieliny i wydaliny przez bezpośredni kontakt. Dlatego też, substancje aktywujące VNO są zazwyczaj przenoszone do jamy tego narządu w formie ciekłej. Odbywa się to poprzez tzw. mechanizm pompujący, związany ze zmianami stopnia ukrwienia. Tkanka jamista po napełnieniu krwią powoduje wypływ płynu z jamy VNO. Natomiast zwężenie ścian naczyń żylnych prowadzi do wzrostu objętości jamy i wytworzenia podciśnienia, które umożliwia zasysanie płynu do wnętrza jamy narządu. Istotną rolę spełniają tutaj unerwienie oraz obecność komórek mięśni gładkich ułożonych promieniście wokół ściany zatoki żylnej w tkance jamistej. W przenoszeniu feromonów w narządzie przylemieszowym uczestniczą proteiny lipokalinowe. Lipokaliny mają zdolność wiązania feromonów w fazie ciekłej i transportowania ich do swoistych receptorów obecnych na powierzchni neuronów VNO.
Komórki receptorowe stanowią dwubiegunowe neurony. Dendryty tych komórek zakończone są pęcherzykami pokrytymi mikrokosmkami, które wystają nad powierzchnią nabłonka. Natomiast ich aksony przewodzą impulsy od perikarionów do dodatkowej opuszki węchowej (opuszki lemieszowo-nosowej).
Związki semiochemiczne są ligandami dla receptorów, wśród których można wyodrębnić dwie grupy (V1R i V2R). Receptory V1R ulegają ekspresji w komórkach o krótkich dendrytach, które wysyłają projekcje do głowowej części dodatkowej opuszki węcho wej. Drugą grupę stanowią receptory V2R, obecne w neuronach z długimi dendrytami, przekazującymi impulsy do ogonowej części opuszki lemieszowo-nosowej. Obie subpopulacje (V1R i V2R), podobnie jak receptory węchowe u owadów, należą do receptorów związanych z białkami G. Przyłączenie liganda (cząsteczki feromonu) do receptorów V1R lub V2R powoduje aktywację błonowego enzymu, fosfolipazy C, która katalizuje powstawanie trifosforanu inozytolu (IP3), co doprowadza do podwyższenia stężenia jonów Ca2+ w komórce. Skutkuje to uruchomieniem wielu szlaków wewnątrzkomórkowych prowadzących m.in. do zmian właściwości błony komórkowej generując potencjały czynnościowe.
Powstałe w ten sposób impulsy nerwowe są przesyłane do dodatkowej opuszki węchowej poprzez tzw. nerwy lemieszowe, utworzone z aksonów komórek receptorowych. Informacja zostaje przekazana do struktur układu limbicznego z pominięciem ośrodków kory odpowiedzialnych za świadomą percepcję węchową. Układ limbiczny jest najstarszą ewolucyjnie częścią mózgu, która kieruje emocjonalnymi reakcjami oraz wrodzonymi, instynktownymi zachowaniami. Feromony są przekazywane zatem w sposób podświadomy za pośrednictwem VNO, dlatego narząd ten zwany jest często „szóstym zmysłem”.
Narząd lemieszowo-nosowy jest dobrze rozwinięty u większości ssaków, a szczególnie u zwierząt drapieżnych i przeżuwaczy. Ponadto, w przeciwieństwie do ptaków i ryb, organ ten posiadają płazy oraz niektóre gady. VNO wykryto także u człowieka, lecz początkowo uważany był jedynie za cechę atawistyczną. W 1991 r. wykazano jednak, iż określone związki semiochemiczne aktywują ludzki VNO w sposób zależny od płci i odmienny niż główny narząd węchowy. Dowiedziono, że poprzez organ Jacobsona możliwa jest stymulacja pracy gruczołów wydzielania wewnętrznego oraz autonomicznego układu nerwowego. Wynika z tego, iż narząd lemieszowo-nosowy u człowieka jest funkcjonalnym organem o cechach dymorfizmu płciowego. W porównaniu do zwierząt cechuje go jednak szczątkowa budowa. Długość tego narządu wynosi zaledwie 2 do 8 mm, natomiast średnica nie przekracza 1 mm. Brak tkanki jamistej oraz dużych naczyń krwionośnych sugeruje, iż związki semiochemiczne przedostają się do jamy narządu na zasadzie dyfuzji i stymulują go jako substancje lotne.
Obecność VNO wykazano u 46,3% badanych osób dorosłych i stwierdzono, iż narząd ten występuje częściej u mężczyzn niż u kobiet. Wynikałoby z tego, że tylko część populacji ludzkiej potrafi reagować na feromony. Okazało się jednak, iż sygnały semiochemiczne mogą być także odbierane przez nabłonek węchowy zarówno u ludzi, jak i zwierząt, a VNO posiada dodatkowo zdolność percepcji związków nie będących feromonami. U ssaków za odbiór feromonów odpowiedzialny jest zatem cały narząd węchu, a jego składowe, czyli okolica węchowa i narząd lemieszowo-nosowy wzajemnie się dopełniają.
Badacze sklasyfikowali związki o charakterze feromonów biorąc pod uwagę rozmaite cechy tych substancji. Najpopularniejszą klasyfikacją feromonów jest podział ze względu na pełnione funkcje. Biorąc pod uwagę to kryterium, wyróżnia się feromony alarmujące, płciowe, zniechęcające, znaczące, ścieżkowe, agregacyjne, zwiększające sprawność fizyczną zwierzęcia czy redukujące stopień wsobności. W świecie przyrody obserwować można również wielofunkcyjne działanie pewnych związków o charakterze infochemicznym. Substancje o właściwościach feromonów odgrywają szereg istotnych ról zarówno w świecie organizmów prostych, prokariotycznych, jak i eukariotycznych, łącznie z człowiekiem.
Wiele wiadomo o feromonach zwierzęcych, znacznie mniej o feromonach bakteryjnych, grzybowych czy roślinnych, które są niezbędne do przetrwania tych organizmów. W latach 70. XX w. A. S. Khokhlov odkrył czynnik A (lakton butyrylu), substancję chemiczną warunkującą produkcję streptomycyny i sporulację u Streptomyces griseus. Badania te zaowocowały identyfikacją kolejnych cząstek sygnalizacji między komórkami bakteryjnymi. Obecnie badacze podkreślają bardzo istotną rolę komunikacji chemicznej w adaptacji bakterii do warunków środowiska oraz zasiedlaniu nowych nisz. Bakteryjne feromony kontrolują najważniejsze procesy życiowe mikroorganizmów, takie jak: sporulacja, koniugacja, bioluminescencja, podział komórki i wirulencja. Związki semiochemiczne są także obecne w świecie grzybów. Gametangia grzybów z rodzaju Allomyces produkują syreninę, która jest silnym atraktantem płciowym gamet męskich. Jest ona również pierwszym opisanym feromonem płciowym roślin. Ciekawym przykładem feromonów roślinnych są związki wydzielane przez liczne gatunki storczyków zapylanych przez osy i pszczoły. Rośliny te produkują substancje identyczne z atraktantami seksualnymi wytwarzanymi przez samice owadów w celu zwabienia samca. W ten sposób kwiat zostaje zapylony pyłkiem przenoszonym przez przywabionego samca. Takie „oszukiwanie” jest powszechne wśród storczyków – ponad 30% opisanych gatunków to „oszuści”.
Komunikacja chemiczna jest niezwykle ważna u owadów, szczególnie gatunków społecznych. Feromony ścieżkowe, tj. sygnały mające na celu wyznaczenie „ścieżki” wędrowania dla zwierząt tworzących wspólnoty, najpowszechniej spotykane są w świecie owadów, takich jak mrówki czy pszczoły. Droga wyznaczana za pomocą feromonów często prowadzi do źródła pożywienia czy też schronienia. Niezwykle interesującą grupę feromonów stanowią związki dyskryminujące. Feromony dyskryminujące są szczególnie istotne dla funkcjonowania owadzich społeczności, ponieważ zapewniają odpowiednią hierarchię. Pod względem chemicznym są to głównie kwasy karboksylowe. Należy do nich między innymi kwas 9-oksodec-2-enowy — lotny kwas tłuszczowy produkowany przez królową pszczół. Feromon ten hamuje rozwój jajników u pszczół robotnic, a tym samym uniemożliwia rekrutację nowej królowej spośród nich. Związek ten jest także wykorzystywany przez niezapłodnioną królową jako atraktant seksualny.
Feromony płciowe to z pewnością najbardziej znane substancje semiochemiczne. Do tej grupy zalicza się różne związki chemiczne. Mogą to być aldehydy, ketokwasy, węglowodory, kwasy karboksylowe, związki alifatyczne czy aromatyczne. Główną ich funkcją jest udział w procesie reprodukcji. Pomagają one zwierzętom zarówno w wyborze partnera seksualnego, synchronizacji rui, jak i opiece nad potomstwem. Niestety wiedza o nich jest wciąż znikoma.
Ssaki pozostawiają w środowisku ogromną ilość śladów „zapachowych”. Razem z moczem, odchodami, wydzielinami skóry, układu rozrodczego czy specjalnych gruczołów uwalniane są cząsteczki, od prostych pod względem budowy, lotnych związków, po duże białka, które stanowią informację o płci, wieku, stanie zdrowotnym, statusie reprodukcyjnym zwierzęcia. Pozwala to na identyfikację konkretnego osobnika przez inne oraz na zaznaczenie zajmowanego przez zwierzę terytorium. Przykładowo, psy, lisy i wilki za pomocą moczu znaczą obszar przebywania i polowania. Jednak w pewnych sytuacjach ślady te mogą wprowadzić w błąd odbiorcę, ponieważ na ich charakter istotny wpływ może mieć na przykład dieta lub naturalna mikroflora zwierzęcia, które to parametry mogą zafałszować pozostawioną przez osobnika „wizytówkę”.
W przypadku wielu gatunków zwierząt przebywających w ogrodach zoologicznych rozmnażanie stanowi poważny problem. Tego rodzaju trudności spowodowane są w dużej mierze zanikiem zdolności reagowania lub nieprawidłową reakcją na atraktanty płciowe. Natomiast gryzonie i zwierzęta udomowione z reguły nie mają kłopotu z rozrodem w niewoli.
O tym jak istotną rolę w reprodukcji ssaków pełnią lotne feromony świadczy przykład słonia azjatyckiego. Za kojarzenie par u tego gatunku odpowiedzialny jest octan Z-7dodecen-1-ylu. Substancja ta jest wydzielana wraz z moczem, na kilka dni przed osiągnięciem przez samicę okresu płodnego. Gdy taki sygnał dotrze do samca, często bardzo oddalonego od samicy (źródła feromonu), można zaobserwować charakterystyczne jego zachowanie polegające na specyficznych ruchach trąby oraz wywijaniu warg czyli flehmenu. Słoń w ten sposób zapewnia efektywniejsze docieranie atraktanta do VNO. Następnie samiec rozpoczyna podróż w celu odnalezienia partnerki, oczywiście kierując się drogowskazem jakim jest jej feromon. Prace badawcze skupione nad wyjaśnieniem mechanizmów kontrolujących przebieg godów tych zwierząt, w tym zjawiska komunikacji chemicznej między osobnikami, mają ogromne znaczenie ze względu na ochronę gatunku i zwiększenie jego populacji. W przypadku słoni afrykańskich, nadal trwają badania nad identyfikacją związków sygnalnych związanych z rozrodem.
Nie można zapomnieć o feromonach zaangażowanych w rozmnażanie ryb oraz innych zwierząt wodnych. Środowisko wodne nie sprzyja wymianie większości sygnałów ze względu na znaczną gęstość, zanieczyszczenia oraz objętość. Sygnalizacja chemiczna umożliwia zwierzętom wodnym pokonanie tych ograniczeń. Feromony wydzielane przez ryby synchronizują tarło, co zapewnia skuteczne zapłodnienie ikry, a także regulują migrację i zachowania społeczne. Interesujące jest również, że w przypadku niektórych gatunków feromony kontrolują ustalanie hierarchii samców, co jest bardzo ważne podczas godów.
Kolejnej klasyfikacji feromonów na dwie grupy dokonano na podstawie rodzaju efektu jaki wywołują oraz czasu niezbędnego do zapoczątkowania reakcji fizjologicznej. Feromony sygnalizujące (wywoływacze), to substancje powodujące u odbiorcy natychmiastową, łatwo i szybko dostrzegalną reakcję behawioralną. W ten właśnie sposób działają między innymi feromony agregacyjne czy ścieżkowe
Do drugiej grupy zalicza się feromony modyfikujące (inicjujące), które wywołują zmiany fizjologiczne po dłuższym czasie. Obecnie trwają intensywne prace nad wyjaśnieniem mechanizmów ich działania.
Wiele efektów działania feromonów ma swoje nazwy od nazwiska ich odkrywców.
Efekt Whitten’a polega na stymulującym działaniu substancji zapachowej zawartej w moczu samca na cykl rujowy samicy. Wesley Kingston Whitten wykazał, że odizolowane od samców samice myszy i świnki morskiej wchodzą w okres estrus w krótkim czasie po pojawieniu się samca zdolnego do rozrodu. Związek chemiczny zawarty w moczu samca wywołuje i przyspiesza ruję samicy, a w grupie samic przebywających razem doprowadza do synchronizacji rui.
Efekt Vandenbergh’a powoduje przyspieszenie dojrzewania samic hodowanych we wspólnej klatce.
Efekt Bruce’a związany jest bezpośrednio z zaburzeniami ciąży u gryzoni. W okresie tuż po zapłodnieniu, samica, która wyczuła woń samca różniącą się od zapachu jej partnera, wraca do stanu rui, a tym samym nie dochodzi do implantacji zarodka. Dzieje się tak na skutek zaburzeń neuroendokrynnych oraz zaniku produkcji hormonów jajnikowych niezbędnych do wytworzenia odpowiedniego endometrium. Przypuszcza się, iż jest to cecha adaptatywna samców, gdyż dzięki efektowi Bruce’a samica po 1 do 4 dni jest znowu gotowa do zapłodnienia. Korzyści samicy są niejasne. Prawdopodobnie wynikają z potrzeby zabezpieczenia swojego przyszłego potomstwa na wypadek, gdyby obcy samiec chciał je zabić po porodzie. Ponadto samica zmniejsza wydatki energetyczne na reprodukcję. Feromon ten zwiększa też szanse na przekazanie swoich genów przez określonego osobnika, zwłaszcza dominującego samca. Na efekt Bruce’a wpływa czas trwania ekspozycji, doświadczenie seksualne i zachowanie obcego samca. Znane są sposoby przeciwdziałania efektowi Bruce’a, którymi są eksperymentalne podawanie egzogennej prolaktyny, progesteronu czy też blokowanie podwzgórza za pomocą rezerpiny.
Efekt Lee-Boota z kolei, dotyczy zahamowania rui u samic przebywających razem, przy braku samca. Ruja jest ograniczona, często występuje ciąża rzekoma lub okresy spokoju płciowego. Zjawisko to nie zostało jeszcze dokładnie opisane. Przypuszcza się, że może on mieć wpływ na redukcję liczebności populacji o nadmiernym zagęszczeniu.
Efekt Ropartza został odkryty u gryzoni. Polega on na tym, że zapach innych osobników powoduje nadmierną produkcję kortykosteroidów, co zmniejsza zdolności reprodukcyjne zwierzęcia.
Oddziaływania o charakterze feromonalnym pełnią niezwykle istotną rolę również u człowieka. Do określenia oddziaływań fermonów w świecie ludzi przyczyniły się badania prowadzone przez Marthę McClintock, która stwierdziła synchronizację cykli menstruacyjnych u kobiet mieszkających razem. Zjawisko to, znane jako efekt McClintock, jest niezwykle podobne do obserwowanego u zwierząt efektu Whitten’a. Ta sama badaczka prowadząc eksperyment w jednym z klasztorów zaobserwowała efekt podobny do efektu Lee-Boot’a u myszy. W sytuacji, gdy bielizna pościelowa zakonnic została skropiona męskim moczem, ich cykle miesięczne zostały zakłócone oraz zanikła obserwowana wcześniej synchronizacja menstruacji.
U ludzi istotne wydaje się być także wy stepowanie specyficznej, chemicznej komunikacji pomiędzy matką a jej nowo narodzonym dzieckiem. Model McFarlana zakłada, iż noworodek podczas pierwszego karmienia wytwarza feromon markujący, którym oznacza pierś matki i w kolejnych dniach wykrywa ją po zapachu. Natomiast po ok. 6-ciu tygodniach, dodatkową rolę zaczyna odgrywać substancja infochemiczna wydzielana na piersiach matki, która modyfikuje zachowanie noworodka, co określone zostało jako model Russella.
Wreszcie, feromony można również podzielić ze względu na ich właściwości i sposób działania na: lotne, przenoszone na duże odległości, oraz kontaktowe, których aktywność jest dostrzegana tylko z odległości milimetrów czy centymetrów. Wśród tak dużej grupy substancji infochemicznych występują także feromony w postaci związanej z białkiem, niektóre w formie stałej lub półstałej. Ponadto, związki semiochemiczne obserwuje się, jak już wspomniano, w świecie organizmów wodnych, co wskazuje na ich występowanie również w formie płynnej.
Badania nad feromonami zwierzęcymi przyczyniły się do zrozumienia mechanizmów sterujących zachowaniem zwierząt. Zdobytą wiedzę można wykorzystać na przykład w celu ochrony lub przywrócenia biosferze konkretnego gatunku. Feromony znalazły zastosowanie także w wielu działach przemysłu.
Klasycznym przykładem są perfumy, w skład których dawniej wchodziły feromony zwierzęce (na przykład piżmo, wydzielina gruczołów okołodbytniczych piżmaka, woła piżmowego, kaczki piżmowej i wielu innych zwierząt), które obecnie zastępuje się syntetycznymi odpowiednikami oraz feromonami roślinnymi (takimi jak olejek jaśminowy).
Od kilkudziesięciu lat trwają próby wykorzystania feromonów w walce z owadami. Zastąpienie insektycydów feromonami pozwala uniknąć szeregu zagrożeń, które niesie ze sobą stosowanie chemicznych środków owadobójczych. Przede wszystkim działanie feromonu ogranicza się zazwyczaj do jednego konkretnego gatunku, nie wywołuje reakcji u pozostałych owadów, a zwierzęta nie uodparniają się na feromony. Poza tym, stosowanie feromonów jest tańsze niż wykorzystanie insektycydów i nie powoduje zanieczyszczania środowiska. Do odłowu owadów służą tzw. pułapki feromonowe. Jako wabiki stosuje się w nich przeważnie syntetyczne feromony płciowe bądź agregacyjne, w stężeniach większych niż te spotykane w naturze. Owady podążając w kierunku źródła feromonu, wpadają w pułapkę, którą może być taśma posmarowana substancją klejącą i feromonem lub puszka zaopatrzona w wejście uniemożliwiające ponowne wydostanie się na zewnątrz, wypełniona wodą lub środkiem owadobójczym.
Sygnalizacja chemiczna jest jedną z naj starszych form komunikowania się między istotami żywymi. Świat feromonów jest bardzo różnorodny i wciąż kryje mnóstwo tajemnic. Wiele odkryć w tej dziedzinie jest jeszcze przed nami. Badania skoncentrowane na tych związkach zapewne pomogą nam lepiej zrozumieć nasze zachowania i odczucia oraz otaczającą nas przyrodę.
żródło: Barata E. N., huBBarD p. c., alMeiDa o. g., MiranDa a., canário a. V., 2007. Male urine signals social rank in the Mozambique tilapia (Oreochromis mossambicus). BMC Biol. 5, 54.BarBier M., leDerer E., 1960. Study of the secretion of the mandibular glands of the queens and workers of bees (Apis mellifica) by gas phase chromatography. C R Hebd. Seances Acad. Sci. 250, 4467.Baxi K. N., DorrieS k. M., eiSthen H. L., 2006. Is the vomeronasal system really specialized for detecting pheromones? Trends Neurosci. 29, 1–7.Boch R., Shearer D. a., Stone B. C., 1962. Identification of isoamyl acetate as an active component in the sting pheromone of the honey bee. Nature 195, 1018–20 Brennan P. A., kenDrick K. M., 2006. Mammalian social odours: attraction and individual recognition. Phil. Trans. Soc. 361, 2061–2078.Bruce H. M.,1959. An exteroceptive block to pregnancy in the mouse. Nature 184, 105.Wikipedia,ButenanDt A., BeckMann r., StaMM D., hecker e., 1959. Zeitschrift fur Naturforschung 14, 283. cavaggioni A., Mucignat-caretta c., caretta a., 1995. Acceleration of puberty onset in female mice by male urinary proteins. J. Physiol. 486, 517–522.Doving K. B., trotier D., 1998. Structure and function of the vomeronasal organ. J. Exp. Biol. 201, 2913–2925. el-SayeD a. M., Suckling D. M., wearing c. h., ByerS J. A., 2006. Potential of mass trapping for longterm pest management and eradication of invasive species. J. Econ. Entomol. 99, 1550–1564.epple G., Belcher a. M., kuDerling i., Seller u.,Scolnick l., greenFielD K. L., SMith A. B., 1993 Making the sense out of scents: species differences in scent glands, scent marking behaviour, and scent mark composition in the Callitrichidae. [W:] Marmosets and tamarins: systematics, behaviour and ecology. rylanDS A. B. (red.). Oxford University Press, 123–151. ganSwinDt A., raSMuSSen H. B., heiSteMannn M., hoDgeS j. K., 2005. The sexually active states of free-ranging male African elephants (Loxodonta africana): defining musth and non-musth using endocrinology, physical signals, and behavior. Horm. Behav. 47, 83–91.gelez H., FaBre-nyS C., 2004. The “male effect” in sheep and goats: a review of the respective roles of the two olfactory systems. Horm. Behav. 46, 257–271.gelez H., FaBre-nyS C., 2006. Role of the olfactory systems and importance of learning in the ewes’, Wykład (żródło?), Monika M. Antkiewicz, Anna DziurDzia, Alicja Socha, Maria Mika
Pojęcia:
Dymorfizm (gr. dimorphos: dwupostaciowy) – występowanie w obrębie jednego gatunku zwierząt lub roślin dwóch różnych form, odmiennych pod względem wyglądu, budowy i fizjologii.
Dymorfizm płciowy, dwupostaciowość – rodzaj dymorfizmu przejawiający się różnicami w morfologii samicy i samca jednego gatunku. Zjawisko to jest widoczne u wielu grup zwierząt, np.:u ssaków[ wyraża się większymi rozmiarami samców oraz czasem obecnością dodatkowych cech (np. rogi tylko u jednej z płci)
Filogeneza (gr. φυλη – gatunek, ród i γενεσις – pochodzenie) – droga rozwoju rodowego, pochodzenie i zmiany ewolucyjne grupy organizmów, zwykle gatunków.
Inne materiały
 Zatrucie przez narażenie na narkotyki i leki psychodysleptyczne (halucynogenne)gdzie indziej o nieokreślonym zamiarze
Zatrucie przez narażenie na narkotyki i leki psychodysleptyczne (halucynogenne)gdzie indziej o nieokreślonym zamiarze Uraz mięśnia i ścięgna właściwego innego palca na poziomie nadgarstka i ręki
Uraz mięśnia i ścięgna właściwego innego palca na poziomie nadgarstka i ręki Pozbądź się worków pod oczami i cieni pod oczami dzięki TEMU 3 prostym domowym środkom zaradczym
Pozbądź się worków pod oczami i cieni pod oczami dzięki TEMU 3 prostym domowym środkom zaradczym Zapalenie mięśni w przebiegu kiły
Zapalenie mięśni w przebiegu kiły- Zwykle określana jako masa ciała poniżej płodowego.
 Próchnica cementu
Próchnica cementu
- Znakomity
- Bardzo Dobry
- Dobry
- Przeciętny
- Słaby
- Beznadziejny






Więcej
Czy ludzie agresywni powinni być izolowani i co się dzieje z ich mózgiem?
Czy Polsce grozi epidemia Ćpunów?
Ci ludzie są tak głupi a może tak mądrzy albo chorzy?